


La differenziazione sessuale, neurologia della sessualità
- La differenziazione sessuale
- Il ruolo degli ormoni
- Centri neurali della sessualità e dimorfismi sessuali
- Il ruolo dei feromoni
- Le esperienze che si fanno nella prima fase della vita modificano il comportamento materno dell’età evolutiva.
- Dimorfismi sessuali del sistema nervoso centrale umano, identità di genere e orientamento sessuale
- Fisiologia dell’atto sessuale nella donna e nell’uomo
- In Sigmasofia
La differenziazione sessuale
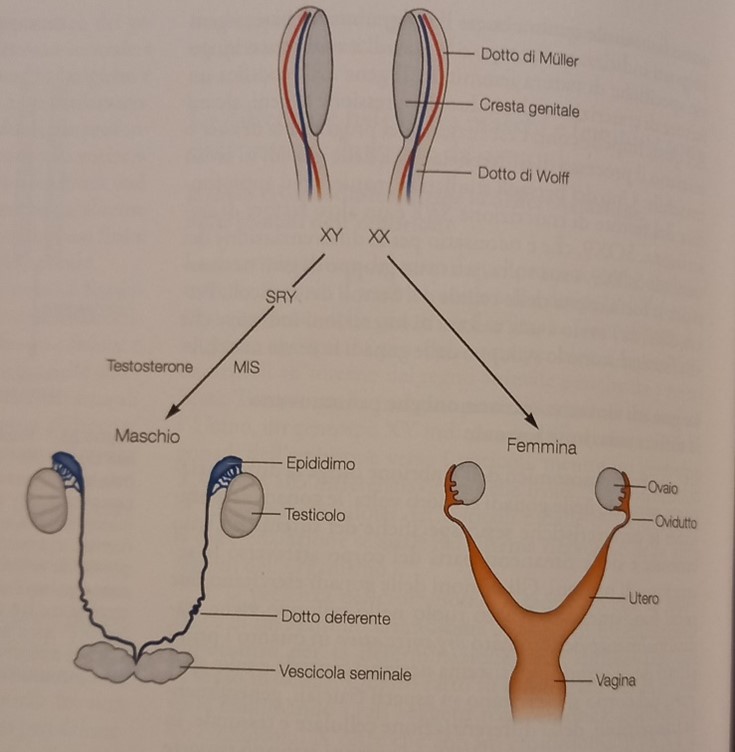
La differenziazione sessuale è determinata dai cromosomi. La specie umana ha 23 coppie di cromosomi, di cui solo una è diversa nei due sessi: le femmine hanno una coppia di cromosomi X (XX), mentre nei maschi il cromosoma X è accoppiato con il cromosoma Y (XY). I cromosomi danno il via alla differenziazione degli organi sessuali maschili e femminili (le gonadi) e la produzione degli ormoni steroidei, prodotti dalle gonadi, dirige la differenziazione sessuale del sistema nervoso centrale e delle altre regioni dell’organismo.
La presenza del cromosoma Y (piuttosto che la mancanza del cromosoma X) determina il sesso maschile. Di base il programma genetico è caratterizzato dalla coppia di cromosomi XX, il gene SRY (sex determining region) codifica l’attività del cromosoma Y. L’inattivazione o la delezione di questo gene comportano l’inversione completa del sesso: i soggetti sono maschi dal punto di vista cromosomico (XY), ma anatomicamente sono femmine, in altri rari casi in cui c’è una mutazione del gene sul cromosoma X gli individui sono cromosomicamente femmine (XX), ma anatomicamente maschi, sterili, quanto molti geni necessari per la funzione degli spermatozoi sono localizzati nel cromosoma Y.
Il gene SRY impedisce l’esecuzione del programma di base (XX), osserviamo come.
Allo stato embrionale, le creste bilaterali genitali (i primordi dei genitali), possiedono entrambe le strutture femminili (i dotti di Müller) e maschili (i dotti di Wolff) che possono differenziarsi in ovaie (i primi) e in testicoli (i secondi).
Negli embrioni XY, il gene SRY a livello delle creste genitali bilaterali induce la differenziazione di questo tessuto in testicoli e dei dotti di Wolff nelle altre parti dei genitali interni maschili, mentre i dotti di Müller vengono riassorbiti.
Negli embrioni XX l’assenza del gene SRY fa sì che le creste genitali si sviluppino in ovaie e i dotti di Müller nelle altre parti dei genitali interni femminili.
Il ruolo degli ormoni
Gli ormoni prodotti dalle gonadi in fase embrionale hanno un ruolo organizzativo in quanto determinano gli effetti sul sistema nervoso centrale e sulle altre parti del corpo riguardo la differenziazione cellulare e tissutale, generalmente irreversibili; in un secondo tempo alcuni di questi ormoni determinano risposte fisiologiche e comportamentali, reversibili, hanno quindi anche un ruolo attivatore.
A livello embrionale, nei maschi, i testicoli in via di sviluppo secernono un ormone di natura proteica inibente il dotto di Müller (MIS –Müllerian inhibiting substance-) e un ormone di natura steroidea: il testosterone. Il MIS provoca la regressione del dotto di Müller, e il testosterone induce il dotto di Wolff a differenziarsi nelle strutture genitali. Nelle femmine, l’assenza del MIS determina la differenziazione delle relative strutture genitali e l’assenza di testosterone determina il riassorbimento del dotto di Wolff.
Gli ormoni steroidei esercitano i loro effetti per tutto l’arco della vita. Questi derivano tutti dal colesterolo. Gli steroidi sessuali si suddividono in androgeni, che promuovono caratteristiche maschili, come il testosterone prodotto dai testicoli, ed estrogeni più progesterone che promuovono caratteristiche femminili: le ovaie producono progesterone e un estrogeno (il 17-β-estradiolo).
La caratteristica sorprendente, che a mio parere sostiene
l’androginia innata
a cui fa riferimento la Sigmasofia, è che oltre al fatto che le creste genitali embrionali possono potenzialmente esprimere entrambi i sessi (prima dell’intervento del gene SRY),
l’ormone femminile progesterone è il precursore del testosterone (maschile)
e
quest’ultimo lo è dell’ormone femminile 17-β-estradiolo.
L’aromatasi è l’enzima che converte il testosterone in estradiolo, ed è presente nelle ovaie (in misura maggiore), nei testicoli e nel sistema nervoso centrale. Le differenze nell’espressione dell’aromatasi sono la causa del dimorfismo sessuale attraverso la regolazione dei livelli in circolo di testosterone ed estrogeno. L’enzima 5-α-reduttasi converte il testosterone in un altro steroide androgeno: il DIT (5-α-diidrotestosterone), a livello di vari tessuti, inclusi i genitali esterni. È responsabile dell’induzione di caratteristiche maschili secondarie, quali i peli e la crescita della prostata, successivamente diventa il responsabile della calvizie.
Ne consegue che un’alterazione del sistema enzimatico a livello ormonale provoca conseguenze di vasta portata.
Esempi di conseguenze dell’alterazione del sistema enzimatico.
L’iperplasia congenita del surrene (CAH –congenital adrenal hyperplasia-)
È una malattia genetica caratterizzata da un’iper produzione di testosterone e dei relativi androgeni a livello del surrene, dovuta a un deficit nella sintesi dei corticosteroidi. Nelle bambine provoca una mascolinizzazione dei genitali esterni (processo detto di virilizzazione). Questa malattia può essere diagnosticata alla nascita e curata mediante intervento chirurgico. Il trattamento con corticosteroidi permette la riduzione dei livelli di testosterone consentendo alle pazienti femmine di avere la pubertà ed essere fertili.
Il deficit di 5-α-reduttasi
Nei feti maschi, questo deficit provoca alla nascita genitali poco virilizzati o nettamente femminili. Questi soggetti cromosomicamente maschi vengono quindi erroneamente fatti crescere come se fossero femmine fine alla pubertà, quando il notevole aumento di testosterone provoca la virilizzazione di tutti i tessuti.
La sindrome da insensibilità completa agli androgeni (CAIS -complete androgen insensitivity syndrome).
Questi pazienti hanno un cromosoma XY ma non hanno la funzionalità del recettore degli androgeni legato a X che abolisce le risposte al testosterone e al DIT, ma poiché il gene SRY è funzionante vi è la formazione dei testicoli ma i dotti di Wolff non si sviluppano non consentendo la discesa dei testicoli e determinando la femminilizzazione dei genitali esterni. Nell’età adulta la maggior parte di questi pazienti opta per la rimozione chirurgica dei testicoli e l’assunzione di ormoni femminili.
Centri neurali della sessualità e dimorfismi sessuali
Il nucleo spinale del muscolo bulbo cavernoso (SNB)
In molti mammiferi, compreso l’Uomo, il midollo spinale contiene un centro sessualmente dimorfico: il nucleo spinale del muscolo bulbo cavernoso (SNB) che ha un ruolo importante nel mantenimento dell’erezione del pene e della fuoriuscita dello sperma nei maschi, dell’erezione del clitoride e delle contrazioni della vagina nelle femmine. Questo muscolo è più grande nei maschi che nelle femmine. Nei ratti questo nucleo contiene molti più neuroni che nelle femmine e ha arborizzazioni dendritiche più estese dovute alla maggiore presenza di testosterone. I motoneuroni di questo nucleo innervano anche il muscolo elevatore dell’ano, implicato nella copulazione, anch’esso più grande nei maschi che nelle femmine. Questo circuito neurale alla nascita non è sessualmente dimorfico, nelle femmine molte fibre muscolari dei muscoli bulbo cavernoso ed elevatore dell’ano, così come molti motoneuroni dell’SNB vanno incontro a morte nella vita postnatale, perciò questo dimorfismo è il prodotto non tanto della generazione maschio-specifica cellulare, quanto della morte femmina specifica delle cellule.
L’ipotalamo
L’ipotalamo, attraverso la regione preottica (anteriore) e il nucleo proprio della stria terminale (BNST) ha un significativo ruolo nei comportamenti di accoppiamento. Nei maschi, i neuroni di queste aree sono più numerosi.
L’ipotalamo è una struttura nervosa posta anatomicamente internamente e al centro dell’encefalo, interposta tra il talamo, superiormente, e l’ipofisi, inferiormente. È connessa, oltre alle strutture appena citate, all’amigdala (tramite la stria terminalis della porzione mediana dell’ipotalamo) e al sistema reticolare del tronco encefalico. Poiché è responsabile della produzione di ormoni rilasciati dall’ipofisi, la maggiore ghiandola endocrina, è considerato appartenere sia al sistema nervoso centrale sia a quello endocrino, costituendone il maggior elemento di connessione. In generale, la funzione dell’ipotalamo è quella di mantenere l’omeostasi corporea[1], ossia una condizione di equilibrio sia per quanto concerne la sfera biochimica e fisica sia per quanto riguarda la sfera comportamentale. L’ipotalamo, infatti, prende parte alla struttura limbica (la cui funzione è prevalentemente legata alle emozioni) e come altra funzione principale, come accennato, produce neurormoni fondamentali alla vita. È implicato anche nei processi di apprendimento, di memoria e altri processi. Tra gli ormoni che produce vi sono le gonadotropine (che regolano le attività delle gonadi maschili e femminili), le corticotropine (che regolano lo stress psichico e fisico), le tireotropine (implicate nel funzionamento della tiroide). Gli ormoni ossitocina e vasopressina sono da esso prodotti ma rilasciati dall’ipofisi e che, come approfondirò più avanti, hanno un ruolo nel comportamento affettivo e sessuale.
Il ruolo dei feromoni
Negli animali, il senso dell’olfatto è necessario per assolvere a funzioni di sopravvivenza (procacciarsi cibo, evitare i predatori ecc.) e i feromoni per influenzare il comportamento di un membro della stessa specie. Studi sui roditori hanno evidenziato che la rimozione del bulbo olfattivo abolisce il comportamento di accoppiamento e i comportamenti aggressivi, ciò indica il ruolo essenziale della stimolazione olfattiva.
Nell’Uomo, l’azione dei feromoni è più controversa, anche se ci sono evidenze di correlazione tra feromoni e funzione sessuale, nonostante l’organo di senso preposto, pur essendo il più antico, è meno sviluppato che in altri mammiferi.
Le esperienze che si fanno nella prima fase della vita modificano il comportamento materno dell’età evolutiva.
L’ipotalamo (l’area preottica e la stria terminale) è importante anche per il comportamento materno nei roditori: la lesione chirurgica di questi centri abolisce il comportamento materno della buona madre (costruire il nido per la prole, tenerli al caldo con il proprio corpo, riportarli al nido se si allontanano). Gli studi condotti in laboratorio sul Ratto hanno evidenziato che le differenze comportamentali esistenti nelle femmine esercitano effetti sulla prole per tutto il resto della vita. È stato riscontrato che alcune madri leccano e governano frequentemente la prole (denominate madri con alto LG), altre lo fanno meno frequentemente (madri con basso LG). Le figlie di madri con alto LG, quando diventano a loro volta madri, presentano un’elevata attività LG e, in condizioni stressanti, sembrano meno ansiose delle figlie di madri con basso LG che, peraltro, quando diventano a loro volta madri, esprimono una bassa attività LG. Ciò sembra evidenziare che la capacità di accudimento della prole e le risposte allo stress siano determinate geneticamente, tuttavia altri studi hanno dimostrato che quando i cuccioli femmina di ratto vengono trasferite a madri adottive, i comportamenti materni che trasferiscono a loro volta sono più simili alla madre adottiva che a quella biologica, ciò evidenzia quindi che l’esperienza durante l’infanzia può determinare caratteristiche comportamentali che perdurano durante la vita ed essere mantenute per molte generazioni.
Tali variazioni sono dovute a modificazioni genomiche e al ruolo degli ormoni.
Una modificazione genomica è basata sull’azione dei glucocorticoidi (ormone steroideo, derivato del cortisolo implicato nella risposta allo stress): la stimolazione tattile provoca l’attivazione del gene recettore dei glucocorticoidi che provoca una riduzione della liberazione degli ormoni ipotalamici implicati nella risposta allo stress, determinandone la riduzione. Un’altra modifica dell’enzima DNA metiltransferasi provoca l’inattivazione del gene del recettore dei glucocorticoidi. Questi processi sono esempi di come una modificazione epigenetica possa determinare l’attivazione o l’inibizione dei geni, provocando in questi animali risposte comportamentali smorzate allo stress.
Un ruolo di fondamentale importanza implicato nelle variazioni comportamentali è svolto dagli ormoni ossitocina e vasopressina. Il primo regola la disponibilità di latte materno[2] e insieme alla vasopressina regolano il legame affettivo della madre e di altri comportamenti sociali. È stato riscontrato che la stimolazione tattile innalza i livelli dell’ossitocina nella prole femminile allevata da madri con alto LG rispetto a quella accudita da madri con basso livello LG. Due recenti osservazioni suggeriscono che questi esperimenti sono validi anche per il comportamento umano: è stato riscontrato che i bambini allevati per lungo periodo in orfanotrofio con modeste attenzioni affettive presentano deficit di diversi comportamenti sociali, e livelli di ossitocina e vasopressina più bassi di quelli allevati da genitori biologici. Bambini che hanno subito abusi spesso non riescono a svolgere bene i loro compiti genitoriali e analisi condotte dopo la morte di adulti, che da bambini avevano subito abusi, rilevano un livello più basso della metilazione dei geni del recettore dei glucocorticosteroidi.
Dimorfismi sessuali del sistema nervoso centrale umano, identità di genere e orientamento sessuale
Seppure esistano aree dimorfiche del sistema nervoso femminile e maschile non sono ancora chiari i meccanismi che danno origine a questi dimorfismi e le relazioni che intercorrono tra essi e il comportamento. I casi considerati clinici relativi al deficit di 5-α-reduttasi e CAIS, poiché in seguito alla pubertà decidono di optare i primi prevalentemente di adottare il genere maschile e i secondi per il genere femminile, suggerisce l’importanza del ruolo ormonale (nei casi di deficit di 5-α-reduttasi l’aumento del livello del testosterone in età puberale a livello cerebrale potrebbe spiegare la scelta di identità di genere maschile, così come la scelta d’identità di genere femminile dei casi di CAIS potrebbe essere dovuta alla mancanza dei recettori degli androgeni), ma non si può escludere l’importanza dell’influenza dei fattori educativi sociali e culturali. Così come nei casi di CAH, c’è chi identifica il proprio genere come maschile, coerentemente alla variazione di genere, altri come femminile con orientamento omo o bisessuale, probabilmente in relazione alla precoce esposizione agli androgeni.
Studi sulla biologia dell’orientamento sessuale umano hanno messo in evidenza le diverse risposte ai feromoni. Questi sono l’AND (androstadione), un metabolita odoroso degli androgeni, presente nel sudore a concentrazione 10 volte più elevate che nelle femmine e l’EST (estradiolo) un metabolita odoroso degli estrogeni, presente nell’urina delle donne gravide. Entrambi possono indurre stimoli sessuali, l’AND per le femmine e l’EST per i maschi, anche a concentrazioni così basse da essere percepite inconsciamente. Le aree attivate da questi feromoni sono state individuate mediante PET (tomografia a emissione di positroni), e sono nuclei (ipotalamici) attivati o inattivi a seconda che appartengano a maschi o femmine eterosessuali (l’AND attiva nuclei nelle femmine eterosessuali che sono inattivi negli uomini eterosessuali e viceversa per l’EST). Negli omosessuali queste arre appaiono invertite (nei maschi omosessuali si attiva il nucleo relativo all’AND e nelle femmine omosessuali quello relativo all’EST). Ciò sembra supportare che il cervello degli omosessuali uomini sia simile a quello delle donne eterosessuali e quello delle donne omosessuali simile a quello degli uomini eterosessuali. Tuttavia a parte i risultati conseguiti sugli studi relativi alle rare sindromi citate, i tentativi di trovare le basi genetiche dell’orientamento sessuale o dell’identità di genere non hanno prodotto risultati attendibili.
Fisiologia dell’atto sessuale nella donna e nell’uomo
I pionieri degli studi scientifici sull’attività sessuale dell’uomo e della donna sono stati il ginecologo e sessuologo statunitense William Masters e la sessuologa statunitense Virginia Johnson negli anni sessanta, che esaminarono, nel corso di 11 anni, oltre diecimila atti sessuali compiuti da circa 700 volontari tra i 18 e gli 89 anni. L’esito dei loro studi fu pubblicato nel 1966 nel volume: L’atto sessuale nell’uomo e nella donna, che contribuì notevolmente a determinare la cosiddetta rivoluzione sessuale, in quanto veniva scardinato il tabù della sessualità e in particolar modo di quella femminile, trattata parimenti a quella maschile. Questi studi suscitarono polemiche e discussioni in tutto il mondo, perché si basavano sull’osservazione diretta del coito e dell’autoerotismo dei volontari, rispetto a precedenti ricerche basate soltanto su interviste e questionari sottoposti agli interessati.
I dati clinici venivano registrati attraverso un poligrafo ed erano inerenti al battito cardiaco, all’attività cerebrale e al metabolismo. Vennero creati anche degli appositi strumenti (i precursori dei sex toys) come il fallo mobile in plexiglas con telecamera incorporata, chiamato “Ulisse”.
Vennero identificate quattro fasi dell’atto sessuale denominate:
- eccitamento
- plateaux
- orgasmo
- risoluzione
Sia l’intensità sia la durata naturalmente possono variare notevolmente da un soggetto all’altro per l’importanza della componente psicologica (che può stimolare, deprimere o inibire una buona risposta sessuale).
Il concetto didesiderio fu successivamente aggiunto da Kaplan (1979) come uno stadio distinto e precedente le fasi indicate, determinando, in caso di disfunzione, la corrispettiva diagnosi di disturbo del desiderio sessuale. Secondo Kaplan inoltre tutte le fasi dell’atto sessuale sono distinte e ognuna può presentare un relativo disturbo.
L’eccitamento
È preceduta da una stimolazione che può essere sia esterna, tattile o visiva, sia interna, dovuta a fantasie erotiche.
Di seguito le diverse manifestazioni
| Nell’uomo | Nella donna |
| • Erezione del pene • Ingrossamento della pelle dello scroto • Ingrossamento dei testicoli • Eventuale erezione dei capezzoli • Aumento del battito cardiaco | • Lubrificazione vaginale • Erezione del glande e del clitoride • Ingrossamento e separazione delle grandi e piccole labbra • Innalzamento dell’utero • Modificazioni delle pareti vaginali • Ingrossamento del seno • Erezione dei capezzoli • Aumento del battito cardiaco |
Fase di Plateau
È la fase che precede l’orgasmo, è quindi una fase di forte intensificazione dell’eccitazione.
| Nell’uomo | Nella donna |
| • dilatazione del pene per il maggior afflusso di sangue fino ai massimi livelli, con colorazione più scura • Innalzamento dei testicoli | • La terza parte più esterna della parete vaginale si ingrossa • L’utero si ingrossa • Il corpo del clitoride si rimpiccolisce e il glande si alza |
Orgasmo
È considerato l’apice del piacere, con una temporanea perdita di contatto con la realtà.
| Nell’uomo | Nella donna |
| • Chiusura dello sfintere della vescica urinaria • Contrazioni ritmiche dei condotti seminali, dell’uretra e della prostata che portano all’eiaculazione di sperma ad intervalli regolari | • Aumento di tensione • Contrazioni della terza parte esterna della vagina • Contrazioni della muscolatura perineale • Contrazioni dell’utero |
Risoluzione
È caratterizzata da uno stato di ritorno iniziale di non eccitazione, nell’uomo coincide con il periodo refrattario, che si manifesta con detumescenza del pene e completa insensibilità a qualsiasi tipo di stimolazione. Nella donna invece non esiste questo periodo e potenzialmente è subito pronta a raggiungere un altro orgasmo.
In Sigmasofia
In Sigmasofia, la Sessualità della Conoscenza permette un vissuto Io-somato-autopoietico notevolmente più esteso, differendo nella pratica su alcuni punti succitati.
Vedi Sessualità della Conoscenza
Note
[1] Regola il senso di sazietà e di sete, la temperatura corporea e la sudorazione, la pressione sanguigna e la frequenza cardiaca, il ritmo sonno-veglia.
[2] L’ossitocina viene prodotta dall’ipotalamo e rilasciata dall’ipofisi, inducendo la contrazione della muscolatura liscia della ghiandola mammaria con conseguente eiezione di latte. L’allattamento a sua volta fornisce uno stimolo sensoriale che viene convogliato all’ipotalamo attraverso vie afferenti. Questo circuito permette il controllo dell’allattamento.
aggredior amigdala autorigenerazione blocchi emozionali cellule gliali cervello comportamento congiungersi contro-dipendenza controdipendenza corpo coscienza differenziazione sessuale dimorfismi dipendenza dormire ecoa emozione emozioni fase rem filosofia fisiologia dell'atto sessuale funzioni vitali istinto neurone neuroni neuroscienze ormoni parte profonda percorsi neuronali processi cognitivi psicosomatica riabilitazione ricarica energetica sensorialità sentimenti sessualità sigmasofia sintomi maestri sistema immunitario sistema nervoso sistema nervoso autonomo sistema somato-sensoriale sonno stimoli corporei
Lascia un commento